Według definicji, poczęcie, zwane też zapłodnieniem, to połączenie dwóch pakietów rekombinowanych genów, z których jeden pochodzi od matki, drugi zaś od ojca. I tutaj można dopatrzyć się trzech postaw, wyróżniających zainteresowanych. Jedni w ogóle się tym nie interesują: - Co? Jak się robi dzieci? Przecież każdy głupi to wie! - Drudzy z grubsza wiedzą, o co chodzi, ale tylko z grubsza, czyli nic nie wiedzą. Trzecia grupa to ci, którzy też nic nie wiedzą, ale chcieliby się dowiedzieć, jak to naprawdę jest, jaki wpływ na potomków będą miały geny przekazane przez nich, no i, co najważniejsze, jaki oni sami mają wpływ na owe geny. By tego dociec, należy wpierw prześledzić proces rekombinacji i wydzielenia owej puli genów, która tylko albo aż w połowie zadecyduje o tym, czy potomek będzie przeciętniakiem, albo będzie wybitnie przystosowany do życia, albo w ogóle do życia zdolny nie będzie.
Pierwotnie znany był jako kwas dezoksyrybonukleinowy, ale z biegiem czasu gdzieś zapodziała się jedna literka i obecnie jest to kwas deoksyrybonukleinowy, co i tak niczego nie zmienia. Ta pokrętna nazwa nie pozostawia wątpliwości, że wymyślili ją chemicy. Strasznie mądrzy chemicy, przynajmniej pragnący za takich uchodzić, z tego też względu nigdy nie wyrażają się w sposób ogólnie zrozumiały, lecz posługują się hermetycznym żargonem zawodowym, czyli taką środowiskową gwarą, niezrozumiałą dla niezwiązanych z tym środowiskiem, a więc przeciętnych, chciałoby się powiedzieć: normalnych ludzi. Przypomina to średniowiecznych medyków, którzy w obecności pacjenta używali tylko łaciny, przez co sprawiali wrażenie, że wiedzą więcej, niż wiedzą.
Przeciętny człowiek pod pojęciem kwas rozumie po pierwsze, że jest to jakiś płyn, a po drugie, że jest kwaśny. W przypadku kwasu deoksyrybonukleinowego ani pierwsze, ani drugie nie jest prawdą, bowiem ani płynny, ani kwaśny on nie jest.
Fakt, zawziąłem się na ten kwas nie-kwas deoksycośtam, ale nie bez powodu. Jako autor publikacji popularnonaukowych zajmuję się tłumaczeniem naukowego bełkotu na ogólnie zrozumiały język ludzki, w związku z czym staram się nie używać naukowych nazw tam, gdzie nie jest to absolutnie konieczne. Tutaj nie widzę nie tylko konieczności, ale choćby potrzeby używania tej, delikatnie mówiąc, dziwnej nazwy, z tego też względu w dalszej części tej publikacji będę używał nazwy prostej i oczywistej: cząsteczka DNA. Krótko i zrozumiale, na ile to oczywiście możliwe.
Podstawowym składnikiem cząsteczki DNA są nukleotydy. Gwoli formalności, ale także, aby dać jako takie wyobrażenie o stopniu skomplikowania, wspomnę tylko, że nukleotydy zbudowane są z reszty cukrowej - pentozy, co najmniej jednej reszty fosforanowej i co najmniej jednej z zasad azotowych: purynowej, pirymidynowej lub flawinowej. Tę informację można szybko zapomnieć i wyobrazić sobie nukleotydy jako kulki.
Gdyby nukleotydy przeciętnej ludzkiej komórki somatycznej ułożyć liniowo, utworzyłyby nić o długości około dwóch metrów. To pokazuje, jak bardzo muszą być one upakowane, by zmieścić się w jądrze komórkowym o średnicy około 6 µm, tj. 6 tysięcznych milimetra, przy czym nie mogą być splątane byle jak, gdyż wówczas zawarta w nich informacja genetyczna byłaby nie do odczytania.
By umieścić nukleotydy w jądrze komórki, konieczne jest więc jakieś uporządkowanie. Dlatego nici nukleotydów są pocięte na cząsteczki i skręcone, ale też nie byle jak skręcone, lecz skręcone precyzyjnie, na wzór helisy*.
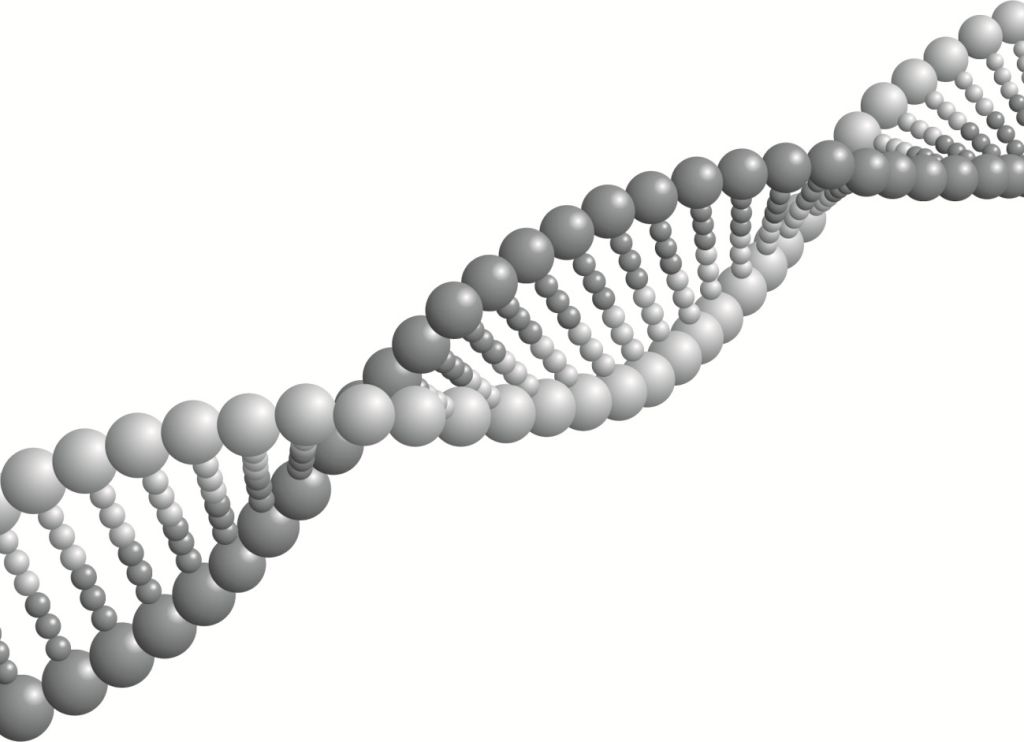
Na rysunku 1 widzimy helisę rozciągniętą, co ma na celu pokazanie sposobu łączenia nukleotydów. W rzeczywistości jest ona ściśle skręcona, tak że przy tym zastosowanym tutaj sposobie prezentacji nukleotydów widzielibyśmy tylko sznurek skręcony z dwóch pasm kulek, wewnątrz wypełniony czymś, czego byśmy nie widzieli. Nie widzielibyśmy zatem, czy i jak obie nici są ze sobą połączone.
Kolejną rzeczą, jaką odkrywamy na rysunku 1, są dwie nici. I tak w rzeczywiści jest, bowiem w skład cząsteczki DNA wchodzą dwie identyczne nici, z tym że biegną one antyrównolegle względem siebie, co znaczy, że koniec jednej znajduje się dokładnie naprzeciwko początku drugiej. Jest to rozwiązanie o tyle praktyczne, że zapisany w nich kod genetyczny z każdej strony można czytać od początku.
W postaci podwójnej helisy cząsteczki DNA przebywają w jądrze komórki, skąd sterują przebiegiem wszystkich jej procesów. W tym stanie komórka funkcjonuje do czasu, aż jakiś tajemniczy impuls, zaprogramowany oczywiście w kodzie DNA, każe jej się rozdwoić.
Podział komórkowy dzieli się na dwa etapy. W pierwszym, zwanym kariokinezą, następuje podział jądra komórkowego, w drugim zaś, zwanym cytokinezą, następuje rozdział cytoplazmy pomiędzy dwie komórki potomne.
* Helisa (linia śrubowa) to krzywa trójwymiarowa zakreślona przez punkt poruszający się ze stałą prędkością po powierzchni walca obracającego się ze stałą prędkością kątową wokół swej osi.
Ze względu na sposób podziału, w ludzkim ciele wyróżniamy dwa typy komórek - somatyczne i płciowe. Różnica między nimi jest taka, że komórka somatyczna dzieli się na dwie identyczne komórki potomne, zaś komórka macierzysta płciowa różnicuje się na cztery komórki, zasadniczo różniące się od komórki wyjściowej, a pod względem genetycznym, o czym będzie potem, także między sobą. Pierwszy z tych podziałów zwie się mitozą, drugi zaś mejozą.
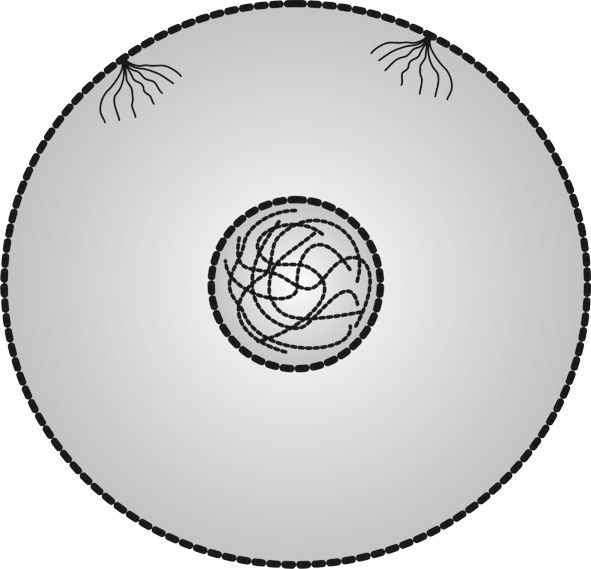 Komórki mają tak różnorodne kształty, że nie sposób wymienić ich wszystkich, z tego względu w rozważaniach teoretycznych zwykło się przedstawiać je jako owalne bryły, mniej lub bardziej zbliżone do kuli. W tym wypadku posłużymy się kulą, mimo że w rzeczywistości tak symetrycznych kształtów komórki nigdy nie mają.
Komórki mają tak różnorodne kształty, że nie sposób wymienić ich wszystkich, z tego względu w rozważaniach teoretycznych zwykło się przedstawiać je jako owalne bryły, mniej lub bardziej zbliżone do kuli. W tym wypadku posłużymy się kulą, mimo że w rzeczywistości tak symetrycznych kształtów komórki nigdy nie mają.
Komórka jest tworem niezwykle skomplikowanym, wypełnionym nieprzeliczoną ilością elementów, z tego też względu w prezentacjach graficznych zwykło się eksponować tylko te elementy, które biorą udział w omawianych funkcjach. Na rysunku 2 wyeksponowane jest jądro komórkowe, z cząsteczkami DNA, oraz dwa wrzeciona podziałowe.
Komórka somatyczna prezentowana na rysunku 2 znajduje się w interfazie, czyli okresie międzypodziałowym. Co istotne, jedynie w interfazie komórka jest - można by rzec - sobą, gdyż pełni funkcje wyznaczone tkance, w skład której wchodzi.
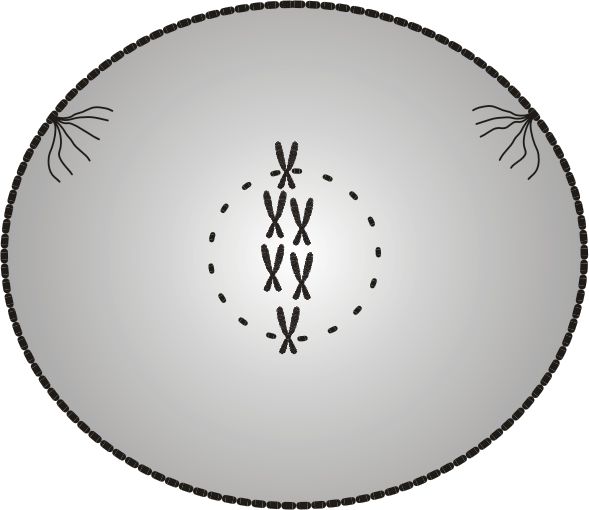
 Profaza to wstępne stadium podziału komórkowego. W fazie tej komórka przestaje pełnić funkcje, które pełniła w interfazie, i przygotowuje się do podziału.
Profaza to wstępne stadium podziału komórkowego. W fazie tej komórka przestaje pełnić funkcje, które pełniła w interfazie, i przygotowuje się do podziału.
Można powiedzieć, że w profazie komórka przestaje być sobą, ponieważ dwuniciowy kod DNA przestaje pełnić w niej jakiekolwiek funkcje, zaś jego rolę przejmuje jednoniciowy kod RNA, zwany kodem pomocniczym.
Za sprawą kodu RNA nici cząsteczek DNA zostają rozłączone, a następnie upakowane w chromatydy, co zapobiega splataniu nici DNA podczas podziału jądra.
Obydwie chromatydy, połączone w punkcie zwanym centromerem, tworzą główny obiekt podziału komórkowego - chromosom.
Oprócz tworzenia chromosomów, w komórce podczas profazy zachodzi szereg innych zmian. Ulega ona spłaszczeniu i uwidoczniają się bieguny, w stronę których przemieszczają się wrzeciona podziałowe. Następuje też zanik osłonki jądra.
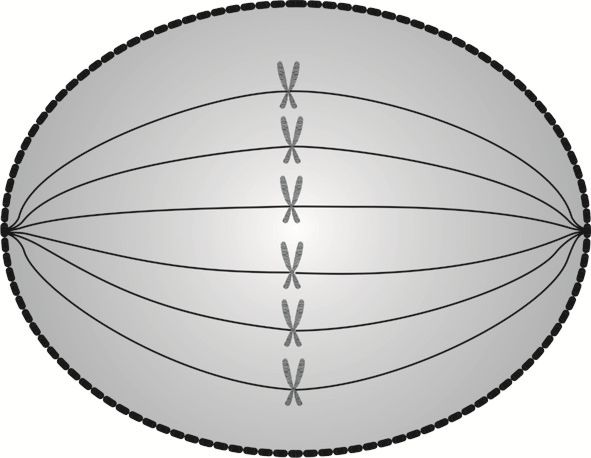 Metafaza jest etapem przygotowującym do rozdziału materiału genetycznego komórki na dwie identyczne, „siostrzane” połówki.
Metafaza jest etapem przygotowującym do rozdziału materiału genetycznego komórki na dwie identyczne, „siostrzane” połówki.
W metafazie wrzeciona podziałowe docierają do biegunów komórki i wykształcają włókna, za pomocą których przyczepiają się do centromerów, czyli punktów złączenia chromatyd.
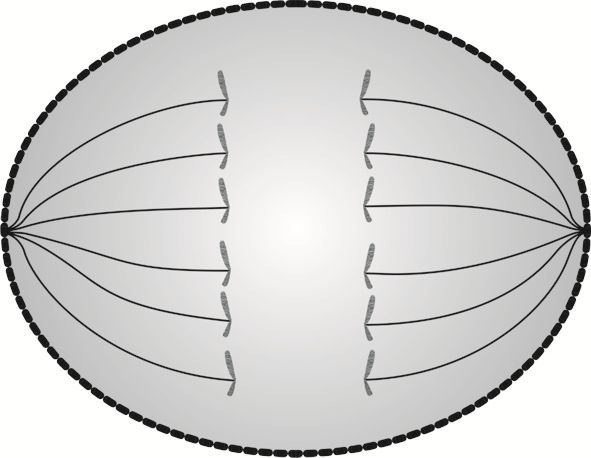 W podziale komórkowym anafaza jest etapem, w którym następuje rozdzielenie chromosomów na dwie identyczne chromatydy siostrzane, zwane odtąd chromosomami potomnymi.
W podziale komórkowym anafaza jest etapem, w którym następuje rozdzielenie chromosomów na dwie identyczne chromatydy siostrzane, zwane odtąd chromosomami potomnymi.
W anafazie włókna wrzecion podziałowych przeciągają chromosomy potomne w kierunku biegunów komórki. Organelle* komórki dzielą się na dwa zespoły takiej samej wielkości i podążają za chromosomami potomnymi.
*Organellum to każda znajdująca się w komórce struktura oddzielona od cytoplazmy błoną, wyspecjalizowana do pełnienia określonej funkcji. Prócz jądra (w interfazie, gdy jest otoczone błoną i pełni swoje funkcje), typowe organelle to: mitochondria, lizosomy, wakuole, siateczka śródplazmatyczna, aparat Golgiego.
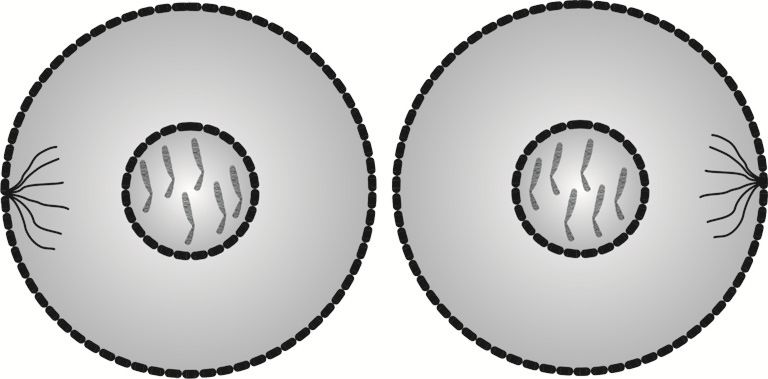
 Telofaza kończy kariokinezę (podział jądra), która jest wstępnym etapem cytokinezy, czyli podziału komórkowego.
Telofaza kończy kariokinezę (podział jądra), która jest wstępnym etapem cytokinezy, czyli podziału komórkowego.
W telofazie następuje zredukowanie włókien wrzecion podziałowych oraz odtworzenie osłonek jąder, w których zachodzą procesy prowadzące do odworzenia cząsteczek DNA.
W telofazie pojawia się w płaszczyźnie równikowej bruzda będąca zapowiedzią ostatecznego podziału komórkowego - cytokinezy.
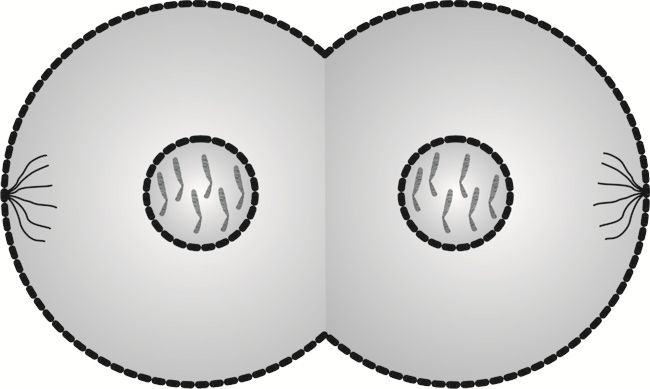
 Cytokineza to ostatni etap podziału komórkowego, skutkiem którego są dwie komórki. W przypadku komórek somatycznych są one identyczne względem siebie i tkanki, w skład której wchodzą.
Cytokineza to ostatni etap podziału komórkowego, skutkiem którego są dwie komórki. W przypadku komórek somatycznych są one identyczne względem siebie i tkanki, w skład której wchodzą.
W cytokinezie następuje rozdzielenie błon komórkowych obu komórek, skutkiem czego powstają dwie niezależne od siebie komórki potomne.
Teraz już pozostaje tylko rozpakowanie chromosomów potomnych, dobudowanie drugiej nici, by cząsteczki DNA przejęły inicjatywę, zaś skutkiem ich aktywności jest podwojenie organelli oraz rozmiarów komórki. Proces podziału komórek somatycznych - od profazy do cytokinezy - trwa niespełna godzinę.
Po zakończeniu procesu podziału komórka somatyczna wchodzi w interfazę, czyli okres międzypodziałowy, który trwa rozmaicie długo, w zależności od obciążenia tkanki, w skład której wchodzi. Po upływie określonego czasu komórka albo ginie, albo dzieli się, by zastąpić sąsiednią komórkę, gdy ta zginie. W ten sposób tkanki naszego ciała bezustannie regenerują się, czyli odnawiają.
Okres interfazy, czyli de facto długość życia komórek ludzkiego ciała, wzbudza nader powszechną ciekawość. By ją zaspokoić, podam kilka przykładów, które wprawdzie nie wyczerpują zagadnienia, ale dają pewne wyobrażenie o tym, jak często cegiełki, z których jesteśmy zbudowani, ulegają zniszczeniu, po czym są wymieniane na nowe. Do najkrócej żyjących bez wątpienia należą komórki nabłonka jelitowego, które regenerują się średnio co pięć dni (jeśli, rzecz jasna, nie są narażone na kontakt z substancjami wybitnie szkodliwymi, co skraca ich i tak już krótki żywot), ale już komórki położone pod nimi, w głębszych warstwach śluzówki, dożywają nawet piętnastu lat. Nieco dłużej, ale też nie za długo, żyją komórki naskórka, które regenerują się co dwa tygodnie, podczas gdy długość życia położonych pod nimi komórek skóry szacowana jest na lat dziesięć. Krwinki czerwone żyją ledwie cztery miesiące, ale limfocyty pamięci immunologicznej nawet dwadzieścia lat. Komórki wątroby żyją rok, a komórki kostne dziesięć lat.
Osobnym zagadnieniem długowieczności jest mózg, który nie stanowi jednolitej tkanki, lecz jest gmatwaniną najprzeróżniejszych, często diametralnie różniących się między sobą tkanek. Badania komórek mózgu, przeprowadzone na materiale pobranym z kory wzrokowej, wykazały, że wszystkie komórki miały taki sam wiek, jak organizm, z którego zostały pobrane, co rodzi domysły, że komórki kory mózgowej nigdy nie regenerują się. Aliści inne badania wykazują, że niektóre komórki kory mózgowej bywają młodsze od organizmu, z którego zostały pobrane, czyli że jednak regenerują się, ale które i jak często - pewności nie ma.
Jak mniemam, jest to jasne i nader oczywiste, jednakże gwoli formalności wspomnę tylko, że powyższe dotyczy jedynie częściowo organizmu dziecka, w którym okres interfazy, czyli długość życia komórek, jest znacznie krótszy, bowiem oprócz podziału komórkowego wymuszonego przebiegającym także w nim nieustannym procesem regeneracji, mamy do czynienia także z przybieraniem masy poszczególnych tkanek, a w rezultacie całego organizmu, co jest bezpośrednim następstwem podziałów komórkowych.
Nie mówi się o tym otwarcie, ale mejoza to w istocie proces produkcji komórek zdefektowanych, pozbawionych, bagatela, połowy materiału genetycznego. Komórki te, siłą rzeczy, w żaden sposób nie są przydatne organizmowi, który je wytworzył. Ich przeznaczeniem jest tylko - czy może aż - podtrzymanie gatunku, czyli przekazanie genów pokoleniom potomnym. Wprawdzie tylko połowy genów, ale w tym wypadku nie ilość ma znaczenie, lecz jakość, od jakości przekazanego materiału genetycznego zależy bowiem cała (jeśli zostanie zachowana) linia rozwojowa potomków - dzieci, ich dzieci i ich dzieci.
Na to, jaki materiał genetyczny zostanie przekazany potomkom, mają wpływ czynniki zarówno subiektywne, a więc zależne od rodziców, jak i obiektywne, czyli od rodziców niezależne - losowe. W tym rozdziale zajmiemy się wyłącznie wpływem losu, a więc przypadku, na jakość puli genów przekazywanych potomkom.
Mejoza zachodzi tylko w komórkach macierzystych płciowych i składa się z dwóch niezależnych podziałów, w przebiegu nieco podobnych do mitozy. Coś jakby dwie następujące po sobie mitozy i tak samo jak mitoza podzielone na fazy. Cały proces jest nieco skomplikowany, niemniej jednak ważne jest, aby go zrozumieć, dlatego przedstawię tylko fakty naświetlające istotę rzeczy, natomiast nieistotne szczegóły tudzież fachowe nazwy, jeśli wymienienie ich nie jest absolutnie konieczne, po prostu pominę. Prawda na tym nie ucierpi, za to czytelnikowi łatwiej będzie zrozumieć przebieg owego tajemniczego procesu, w efekcie którego diploidalny, czyli podwójny zestaw chromosomów, zostaje przetworzony na haploidalny, czyli pojedynczy zestaw chromosomów.
W początku pierwszej profazy mejotycznej, zwanej profazą I, zachodzą zmiany takie same, jak w profazie mitotycznej - nici cząsteczek DNA zostają rozłączone i uformowane w chromatydy, zaś dwie chromatydy tej samej cząsteczki tworzą chromosom. Teraz zaczyna być inaczej niż w mitozie, bowiem w tym etapie mejozy chromosomy łączą się w pary zwane chromosomami homologicznymi.
Dobór chromosomów tworzących chromosomy homologiczne nie jest przypadkowy. Wprost przeciwnie - są one sobie wręcz przypisane, bowiem są to chromosomy niejako bliźniacze - o takiej samej budowie i zawierające takie same geny. Jedyne, co je różni, to to, że jeden pochodzi od ojca, drugi zaś od matki.

Para chromosomów homologicznych składa się z czterech chromatyd. Chromatydy leżące naprzeciwko siebie zwane są siostrzanymi, zaś leżące krzyżowo - niesiostrzanymi. Rozróżnienie to ma istotne znaczenie w objaśnieniu zjawiska zwanego crossing-over.
Crossing-over zachodzi podczas łączenia się chromosomów homologicznych w ściśle do siebie przylegające pary - biwalenty. Zanim obydwa chromosomy nałożą się na siebie, dochodzi do spotkania chromatyd niesiostrzanych. Wówczas stykające się ze sobą fragmenty nici pękają i zamieniają się miejscami.
Wymiana materiału genetycznego ma na celu tzw. zmienność genetyczną, skutkiem której potomek nie jest kopią żadnego z rodziców, lecz posiada osobnicze cechy genetyczne. Czasami lepsze, czasami gorsze, ale zawsze inne. Tutaj widać, jak okrutnie natura traktuje poszczególne jednostki, ani trochę się nimi nie przejmując, jeśli bowiem potomek odziedziczy cechy genetyczne gorsze, to zginie, przez co nastąpi przerwanie owej nieudanej puli genetycznej. Jeśli jednak potomek odziedziczy cechy genetyczne lepsze od rodziców, a więc będzie lepiej przystosowany do środowiska, to linia rozwojowa jego genów zapewni gatunkowi ewolucyjny rozwój.
Nie dotyczy to rzecz jasna gatunku ludzkiego, który w sporym zakresie, nierzadko aż zanadto, podporządkował sobie środowisko. Jednakże właśnie u ludzi najlepiej widać, jak bardzo się różnimy, nie tylko zewnętrznie, ale także charakterem, temperamentem, upodobaniami, zdolnościami, umiejętnościami.
Crossing-over nie jest odrębnym etapem profazy I, lecz zachodzi niejako mimochodem, podczas łączenia chromosomów homologicznych w kompleks zwany biwalentem. No i tym się głównie różni mejoza od mitozy, że w dalszych dwóch podziałach komórkowych uczestniczyć będą nie pojedyncze chromosomy, jak ma to miejsce w mitozie, lecz chromosomy zdublowane, czyli biwalenty.
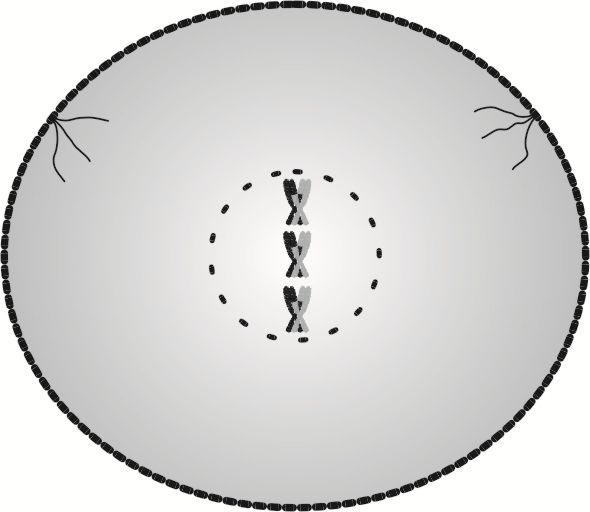 W mejotycznej profazie I, podobnie jak w profazie mitotycznej, następuje lekkie spłaszczenie komórki i zanik osłonki jądra. Wrzeciona podziałowe wędrują ku biegunom, biwalenty ustawiają się w płaszczyźnie równikowej komórki.
W mejotycznej profazie I, podobnie jak w profazie mitotycznej, następuje lekkie spłaszczenie komórki i zanik osłonki jądra. Wrzeciona podziałowe wędrują ku biegunom, biwalenty ustawiają się w płaszczyźnie równikowej komórki.
Jak widzimy na powyższym rysunku, elementów podlegających rozdziałowi na dwie komórki w mejotycznej profazie I jest o połowę mniej, w porównaniu z ilością elementów rozdzielanych w profazie mitotycznej, mimo że ilość chromosomów biorących udział w obydwu fazach podziału komórkowego jest wciąż taka sama - diploidalna. Bierze się to stąd, że w profazie I chromosomy występują po dwa jako biwalenty.
Druga istotna różnica, na którą nie można nie zwrócić uwagi, to ta, że już w profazie I powstają cztery komplety chromosomów, nieco różniących się informacją genetyczną. Różnica ta ma kluczowe znaczenie w dalszym przebiegu mejozy, mającym na celu wytworzenie czterech komórek różniących się nie tylko od wyjściowej macierzystej komórki płciowej, ale różniących się także między sobą.
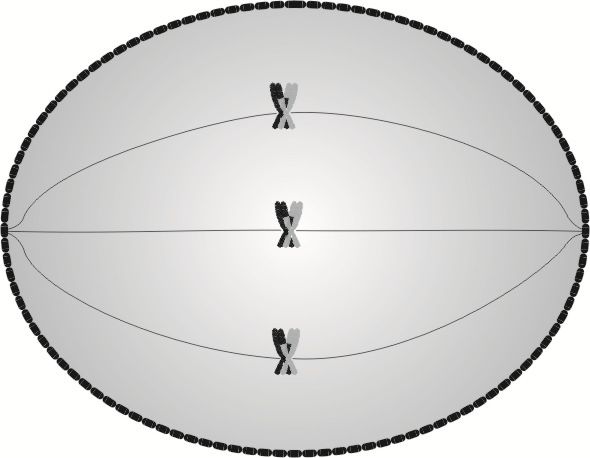 Przebieg mejotycznej metafazy I jest w gruncie rzeczy taki sam, jak przebieg metafazy mitotycznej. Także tutaj wrzeciona podziałowe ustawiają się na biegunach komórki i wykształcają włókna, za pomocą których przyczepiają się do centromerów, z tym że w tym wypadku każde włókno przyczepia się do dwóch centromerów jednocześnie.
Przebieg mejotycznej metafazy I jest w gruncie rzeczy taki sam, jak przebieg metafazy mitotycznej. Także tutaj wrzeciona podziałowe ustawiają się na biegunach komórki i wykształcają włókna, za pomocą których przyczepiają się do centromerów, z tym że w tym wypadku każde włókno przyczepia się do dwóch centromerów jednocześnie.
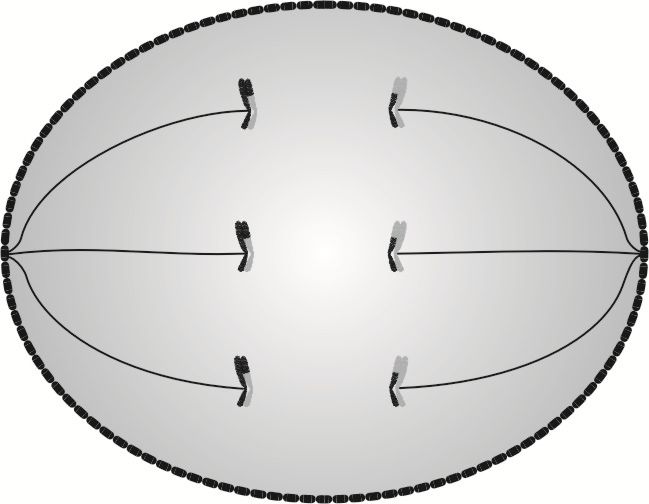 W anafazie I następuje rozdzielenie siostrzanych chromatyd od niesiostrzanych poprzez rozerwanie biwalentów. W tym miejscu należy zaznaczyć, że chromatydy siostrzane nie są już swoim lustrzanym odbiciem, ponieważ różnią je drobne zmiany genetyczne, które zaszły podczas crossing-over. W anafazie I włókna wrzecion podziałowych, skręcając się (czego na rysunku nie widać), rozrywają biwalenty na pary chromatyd siostrzanych, a następnie (wciąż skręcając się) przeciągają je w kierunku przeciwległych biegunów komórki.
W anafazie I następuje rozdzielenie siostrzanych chromatyd od niesiostrzanych poprzez rozerwanie biwalentów. W tym miejscu należy zaznaczyć, że chromatydy siostrzane nie są już swoim lustrzanym odbiciem, ponieważ różnią je drobne zmiany genetyczne, które zaszły podczas crossing-over. W anafazie I włókna wrzecion podziałowych, skręcając się (czego na rysunku nie widać), rozrywają biwalenty na pary chromatyd siostrzanych, a następnie (wciąż skręcając się) przeciągają je w kierunku przeciwległych biegunów komórki.
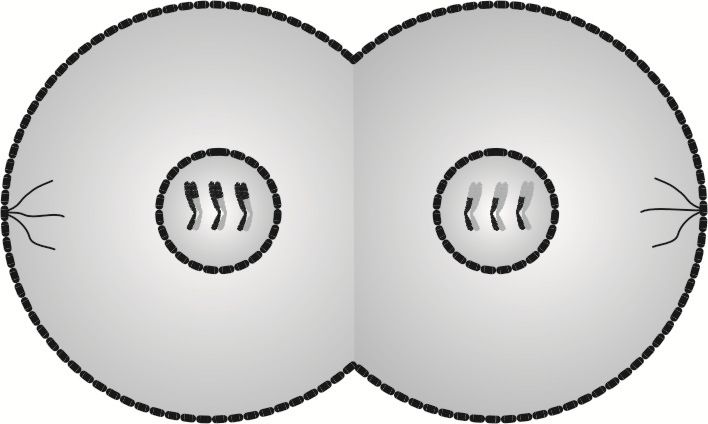 Telofaza I rozpoczyna się, gdy chromatydy siostrzane są już rozdzielone i oddalone od siebie na maksymalną odległość. W telofazie I następuje zredukowanie włókien wrzecion podziałowych oraz odtworzenie osłonek jąder. W płaszczyźnie równikowej komórki pojawia się bruzda podziałowa, będąca zapowiedzią podziału cytoplazmy.
Telofaza I rozpoczyna się, gdy chromatydy siostrzane są już rozdzielone i oddalone od siebie na maksymalną odległość. W telofazie I następuje zredukowanie włókien wrzecion podziałowych oraz odtworzenie osłonek jąder. W płaszczyźnie równikowej komórki pojawia się bruzda podziałowa, będąca zapowiedzią podziału cytoplazmy.
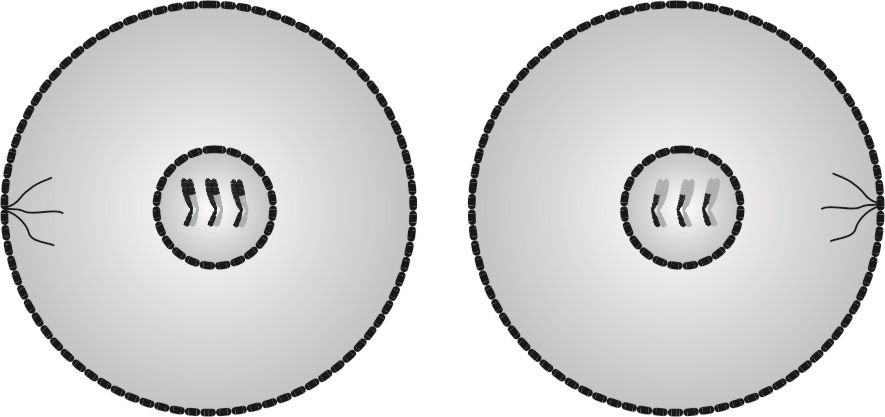 Posługując się językiem technicznym, komórki powstałe w wyniku cytokinezy I to półfabrykaty służące do wyprodukowania produktu ostatecznego. Z tego względu w ich jądrach nie są rozpakowywane nici DNA, by odtworzyć helisę DNA, lecz jako chromatydy siostrzane przechodzą do kolejnego podziału.
Posługując się językiem technicznym, komórki powstałe w wyniku cytokinezy I to półfabrykaty służące do wyprodukowania produktu ostatecznego. Z tego względu w ich jądrach nie są rozpakowywane nici DNA, by odtworzyć helisę DNA, lecz jako chromatydy siostrzane przechodzą do kolejnego podziału.
W cytokinezie I następuje rozdzielenie błon komórkowych obu komórek, więc powstają dwie komórki, różniące się między sobą kodem genetycznym.
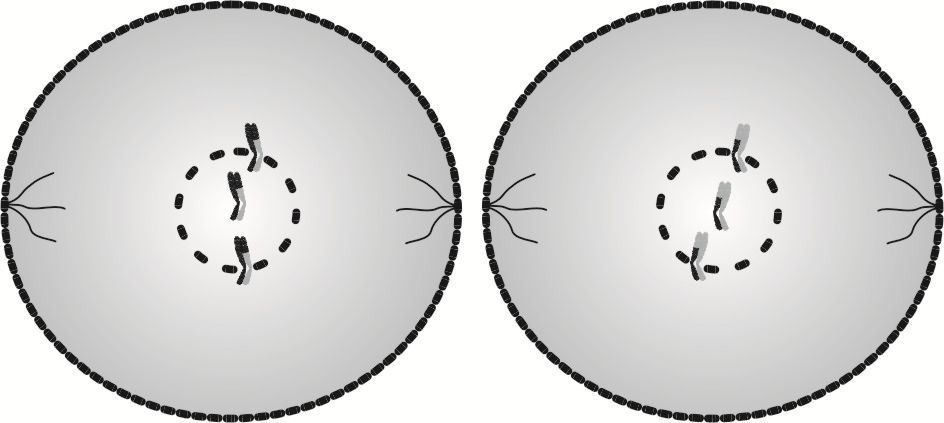 Profaza II rozpoczyna proces drugiego mejotycznego podziału komórkowego. Rozpoczyna się tuż po cytokinezie I i uczestniczą w niej obie komórki jednocześnie.
Profaza II rozpoczyna proces drugiego mejotycznego podziału komórkowego. Rozpoczyna się tuż po cytokinezie I i uczestniczą w niej obie komórki jednocześnie.
W obydwu komórkach uczestniczących w profazie II następuje odtworzenie brakujących wrzecion podziałowych i rozpad otoczek jąder.
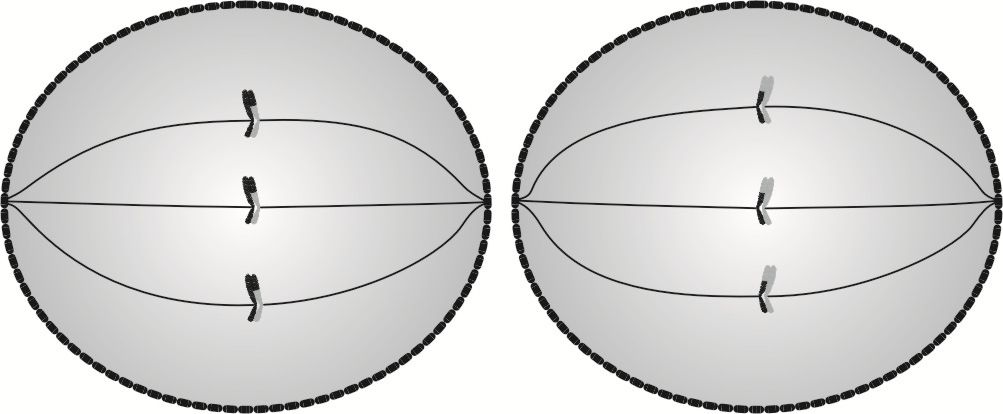 Metafaza II jest wstępem do ostatecznego rozdzielenia chromatyd siostrzanych, które ustawiają się w płaszczyznach równikowych obu komórek, a następnie wrzeciona podziałowe wytwarzają włókna, które przyczepiają się do poszczególnych chromatyd.
Metafaza II jest wstępem do ostatecznego rozdzielenia chromatyd siostrzanych, które ustawiają się w płaszczyznach równikowych obu komórek, a następnie wrzeciona podziałowe wytwarzają włókna, które przyczepiają się do poszczególnych chromatyd.
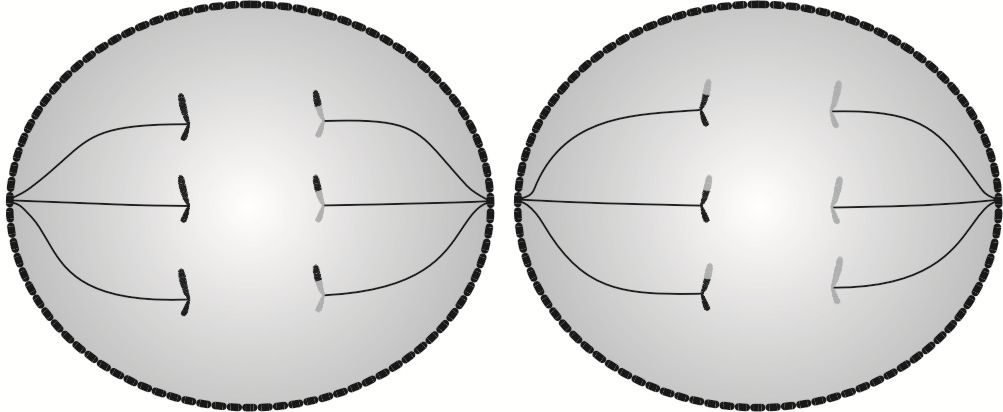 Anafaza II to etap podziału jądra, w którym następuje podzielenie informacji genetycznej na dwoje, skutkiem czego po zakończeniu mejozy komórki będą posiadały haploidalną, czyli o połowę mniejszą liczbę chromosomów, w stosunku do komórki pierwotnej.
Anafaza II to etap podziału jądra, w którym następuje podzielenie informacji genetycznej na dwoje, skutkiem czego po zakończeniu mejozy komórki będą posiadały haploidalną, czyli o połowę mniejszą liczbę chromosomów, w stosunku do komórki pierwotnej.
W anafazie II następuje rozłączenie chromatyd siostrzanych i przesunięcie ich ku przeciwległym biegunom obu komórek.
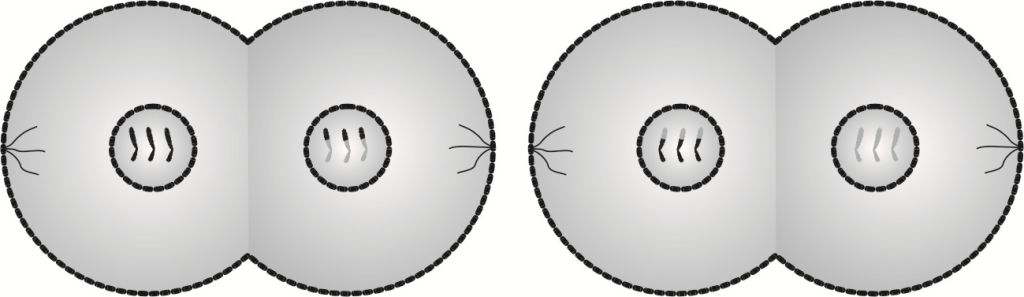 W telofazie II następuje odbudowanie osłonek jąder, a ponadto w płaszczyznach równikowych obu komórek zarysowują się bruzdy podziałowe, będące zapowiedzią rychłego podziału cytoplazmy.
W telofazie II następuje odbudowanie osłonek jąder, a ponadto w płaszczyznach równikowych obu komórek zarysowują się bruzdy podziałowe, będące zapowiedzią rychłego podziału cytoplazmy.
Cytokineza II to faza dojrzewania gamet. Na tym etapie podziału komórkowego zachodzi szereg zmian, zarówno w jądrach komórkowych, jak i w samych komórkach. Nade wszystko zostaje ukończony zapoczątkowany w telofazie II rozdział cytoplazmy obu komórek, wskutek czego z każdej z nich powstają dwie o połowę mniejsze komórki, czyli że produktem końcowym mejozy są cztery komórki.
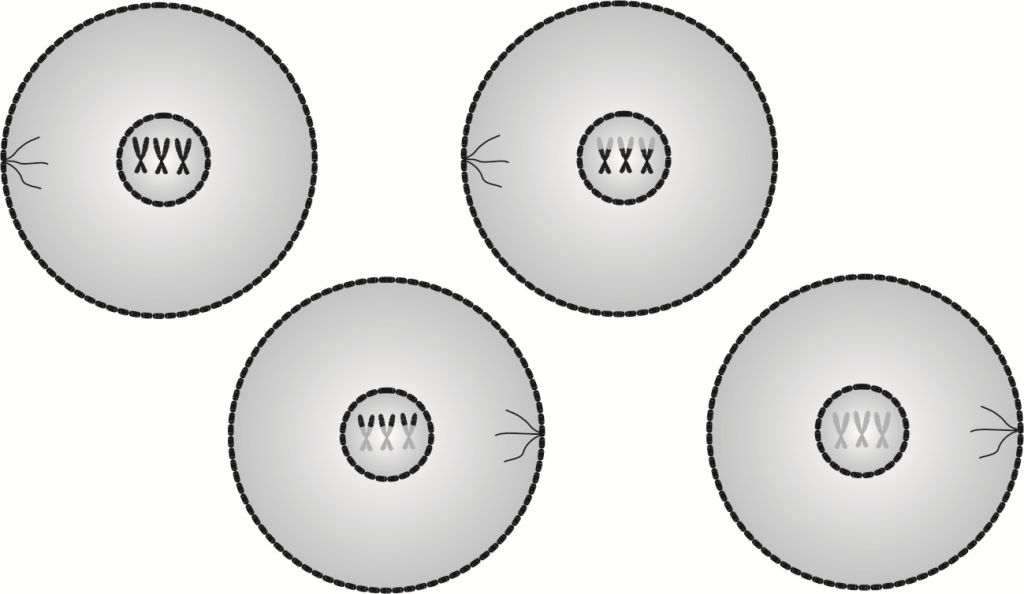
Najistotniejsze zmiany w cytokinezie II zachodzą jednak w jądrach komórkowych. Otóż najpierw każda chromatyda zostaje rozwinięta na jednoniciową DNA, potem zostaje odtworzona druga nić, czyli powstają kompletne cząsteczki DNA, które następnie zostają upakowane w chromosomy. Tak oto w procesie mejozy z jednej macierzystej komórki płciowej powstają cztery gamety, tj. komórki rozrodcze posiadające haploidalną liczbę chromosomów, czyli tylko połowę informacji genetycznej, na dobitkę rekombinowanej w wyniku crossing-over.
Autor: Józef Słonecki